KOSTIMULATORISCHE SIGNALE DER T-ZELLAKTIVIERUNG ALS ANGRIFFSPUNKTE FÜR DIE THERAPEUTISCHE INDUKTION BZW. DURCHBRECHUNG VON TOLERANZ
Die Aktivierung von T-Zellen spielt bei Entzündungsreaktionen und bei Transplantatabstoßungen eine zentrale Rolle. Sie erfordert, dass neben dem Antigen-Erkennungssignal über den T-Zellrezeptor (TZR) kostimulatorische Signale über akzessorische Rezeptoren wie CD2 oder CD28 ausgelöst werden. Antigenerkennung ohne kostimulatorische Signale ist kein neutrales Ereignis für T-Zellen, sondern führt zu einer antigenspezifischen Unreaktivität (Anergie). Dies ist einer der Mechanismen, die dazu führen, dass T-Zellen sich nicht gegen körpereigene Strukturen richten. Intrazelluläre Moleküle, die kostimulatorische Signale vermitteln und deren regulatorischen Enzyme stellen somit interessante Zielstrukturen für neue Strategien zur Immunmodulation dar.
Kostimulatorische Signale werden in vivo durch Bindung akzessorischer Rezeptoren an ihre Liganden (CD80, CD86 und CD58) auf Antigen-präsentierenden Zellen (APZ) ausgelöst. Während der Aktivierung von T-Zellen durch APZ wird an der Kontaktzone eine komplexe Struktur ausgebildet. Analog zu neuronalen Synapsen wurde diese Struktur als reife „Immunologische Synapse“ (IS) bezeichnet (Abb.1). Sie besteht aus einem inneren Aktivierungsbereich, cSMAC (engl. central supramolecular activation cluster), in dem sich der TZR, der akzessorische Rezeptor CD28, sowie die Protein Kinase C θ (PKCθ) anreichern und einem umliegenden Adhäsionsring, pSMAC (engl. peripheral SMAC), in dem LFA-1, CD2 sowie der Membrananker Talin lokalisiert sind. Rezeptoren mit einem großen extrazellulären Bereich, wie die Tyrosin-Phosphatase CD45, befinden sich in einem Bereich außerhalb des pSMACs. Dieser Bereich wird als dSMAC (engl. distal SMAC) bezeichnet. Unklar ist bisher noch, wie die hoch-organisierte Ring-Struktur in der Synapse entsteht. Sicher ist, dass sowohl Kostimulation als auch ein dynamisch funktionierendes Aktin-Zytoskelett dafür notwendig sind.
Abb. 1
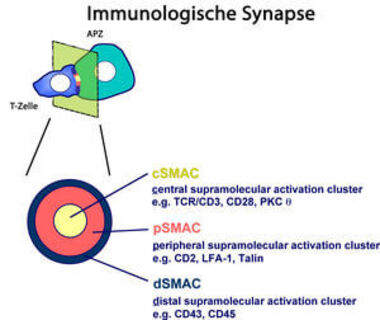
Abb. 1: Die immunologische Synapse
Als Immunologische Synapse (IS) wird die Kontaktzone zwischen der T-Zelle und der Antigenpräsentierenden Zelle (APZ: Dendritische Zelle oder B-Zelle) bezeichnet. In einem Querschnitt durch die Kontaktzone sind in der reifen IS drei in konzentrischen Kreisen aufgebaute Zonen zu unterscheiden. Der innere Kreis (central supramolecular activation cluster (cSMAC)) besteht aus einem zentralen Aktivierungskluster, in dem TZR/CD3, CD28 und PKC theta lokalisiert sind. Der periphere Adhäsionsring (peripheral SMAC oder pSMAC) enthält die Adhäsionsmolekule CD2 und LFA-1. Im äußersten Ring (distal SMAC oder dSMAC) befinden sich große Moleküle, wie CD45 und CD43. Für die Ausbildung dieser Proteinkluster sind sowohl ein intaktes Zytoskelett, als auch T-Zell Kostimulation unabdingbar.
BEDEUTUNG DES AKTIN-BINDENDEN PROTEINS COFILIN FÜR DIE AKTIVIERUNG MENSCHLICHER T-LYMPHOZYTEN
Wir haben kostimulatorische Signalübertragungswege identifiziert, die für die Ausbildung der immunologischen Synapse zwischen T-Zellen und Antigen-präsentierenden Zellen, sowie die T-Zell-Aktivierung essentiell sind. Einer dieser Signalwege führt zur Dephosphorylierung und Aktivierung des Aktin-Zytoskelett-Regulators Cofilin (Wabnitz et al. 2006; Eibert et al. 2004; Lee, K.-H. et al. 2000). Über konfokale Laserscanning-Mikroskopie konnten wir zeigen, dass sich Cofilin im pSMAC der immunologischen Synapse anreichert (Abb. 2). Durch Zerschneiden (Severing) und Depolymerisieren von Aktinfilamenten ermöglicht Cofilin einen dynamischen Umbau des Aktinzytoskeletts (Samstag et al. 2003). Wenn die Aktin-Bindefähigkeit dieses Proteins durch Einschleusen zellpermeabler Peptid-Homologe in vivo blockiert wird, bildet sich keine reife IS aus und die Aktivierung der T-Zellen sowie die Produktion von Zytokinen sind signifikant gestört (Eibert et al. 2004).
ABB. 2
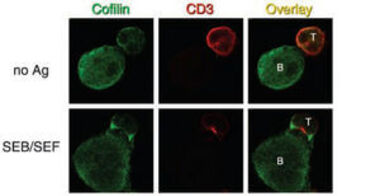
Abb. 2: Cofilin reichert sich im pSMAC der Immunologischen Synapse an
Humane T-Lymphozyten aus dem peripheren Blut (PBT) wurden mit Superantigen-beladenen Raji B-Zellen stimuliert (SEB/SEF) oder mit unbeladenen Raji B-Zellen inkubiert (no Ag). Nach Fixierung wurde Cofilin in grün (Cy2) und CD3 in rot (Cy3) mit entsprechenden Antikörpern angefärbt. Der Overlay zeigt die Überlagerung beider Färbungen. Bei Antigenstimulation (SEB/SEF) reichert sich der antigenspezifische TZR/CD3-Komplex im c-SMAC, Cofilin im p-SMAC an.
Desweiteren haben wir den Signaltransduktionsweg zur Cofilin-Dephosphorylierung charakterisiert (Wabnitz et al. 2006, Samstag et al. 2005; Nebl et al. 2004; Samstag et al. 2003; Ambach et al. 2000). Eine Blockade der Aktivierung oder Expression von Cofilin hemmt die T-Zellwanderung in 3D-Umgebungen (z.B. Gewebe), jedoch nicht die Wanderung auf 2D-Oberflächen (z.B. Endothel), (Klemke et al. 2010). Nun untersuchen wir die Beeinflussung von Cofilin und anderer Proteine durch Veränderungen des Redox-Milieus und deren Auswirkung auf die Kostimulation, Adhäsion und Migration von T-Zellen (zur Übersicht siehe Samstag et al. 2013). Hierdurch hoffen wir, neue Angriffspunkte für die therapeutische Immunmodulation zu identifizieren. Wir konnten zeigen, dass reaktive Sauerstoffspezies in T-Zellen zur Oxidation von Cofilin führen und eine funktionelle Paralyse der T-Zellen auslösen können (Klemke et al. 2008). Lang anhaltender oxidativer Stress führt zur Translokation von oxidiertem Cofilin in die Mitochondrien und induziert dadurch einen programmierten Zelltod (Wabnitz et al. 2010).
BEDEUTUNG DES AKTIN-BÜNDELNDEN PROTEINS L-PLASTIN FÜR DIE AKTIVIERUNG MENSCHLICHER T-LYMPHOZYTEN
Auf der Suche nach Funktionsmolekülen, die selektiv über akzessorische Rezeptoren reguliert werden (s.o.), haben wir ein weiteres interessantes Molekül identifiziert: das Aktin-bündelnde Protein L-Plastin. Über konfokale Laserscanning-Mikroskopie (Abb. 3) und Time-Lapse-Video-Mikroskopie (Abb. 4) konnten wir zeigen, dass L-Plastin innerhalb von Sekunden nach Kontakt mit einer Antigenpräsentierenden Zelle an die Immunologische Synapse verlagert wird (Wabnitz et al. 2007). L-Plastin wird nach Kostimulation am Serin-5 phosphoryliert. Diese Phosphorylierung ist wichtig für die Aktivierung der T-Zellen. Expression einer nicht-phosphorylierbaren Mutante von L-Plastin in untransformierten menschlichen T-Zellen aus dem Blut hat interessanterweise einen dominant negativen (d.h. immunsuppressiven) Effekt auf die T-Zellaktivierung (Wabnitz et al. 2007). Daher könnten L-Plastin und/oder die Kinasen, die für die L-Plastin Phosphorylierung verantwortlich sind, ebenfalls interessante Targets für neue Immunsuppressiva darstellen. In der Tat hemmt das Immunsuppressivum Dexamethason (ein Glucocorticoid) die Phosphorylierung und damit Aktivierung von L-Plastin (Wabnitz et al. 2011).
ABB. 3
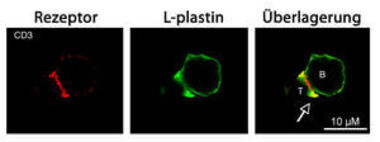
Abb. 3: L-Plastin reichert sich Superantigen-abhängig in den äußeren Bereichen der Kontaktzone zwischen T-Zellen und APZ an
Humane T-Lymphozyten aus dem peripheren Blut (PBT) wurden mit Superantigen-beladenen Raji B-Zellen stimuliert. Nach Fixierung wurde L-Plastin in grün (Cy2) und CD3 in rot (Cy3) mit entsprechenden Antikörpern angefärbt. Die Überlagerung zeigt beide Färbungen. Bei Antigenstimulation (SEB/SEF) reichert sich der antigenspezifische TZR/CD3-Komplex im c-SMAC und L-Plastin im p-SMAC und d-SMAC der Immunologischen Synapse an.
ABB. 4 (VIDEO.AVI)
Abb. 4 (Film): L-Plastin Dynamik in Zell-Kontakten zwischen PBT und Raji B-Zellen
Über Transfektion von cDNA wurde grün fluoreszierendes L-Plastin-EGFP in humanen T-Lymphozyten aus dem peripheren Blut (PBT) exprimiert. Anschließend wurden die PBT mit Superantigen-beladenen Raji B-Zellen (APZ) auf Poly-L-Lysin beschichteten Objektträgern inkubiert. Die Ausbildung der Immunologischen Synapse wurde über Time-Lapse-Videomikroskopie dokumentiert. Für diesen Film wurden im Abstand von 18 Sekunden je ein Digitaler Interferenz Kontrast (DIC, oben) und ein Fluoreszenzbild (unten) aufgenommen. L-Plastin (in Falschfarben dargestellt) verlagert sich innerhalb von Sekunden an die Kontaktstelle mit der APZ.
Eine dynamische Reorganisation des Aktin-Zytoskeletts ist für eine Vielzahl von T-Zellfunktionen wichtig (Samstag et al. 2003). Wie oben beschrieben übernimmt das Aktin-Zytoskelett eine wichtige Funktion bei der Stabilisierung der immunologischen Synapse, d.h. bei der initialen Antigenerkennung. Darüber hinaus ist die Reorganisation des Aktin-Zytoskeletts ein essentieller Bestandteil von Prozessen, die das Zellwachstum (klonale Expansion antigenspezifischer T-Zellen) und die Zellmigration (Homing) steuern. Um die Bedeutung von L-Plastin für die T-Zellmigration und T-Zell-Effektorfunktionen zu klären, wird die Expression von L-Plastin in menschlichen T-Zellen durch siRNA-Ansätze gezielt blockiert. Außerdem werden die Signalübertragungsvorgänge, die die Funktion von L-Plastin steuern, charakterisiert. Wir erhoffen uns, durch Eingriffe in diesen kostimulatorischen Signalweg antigenspezifische Unreaktivität induzieren zu können und das Wachstums- und Migrationsverhalten von T-Zellen zu beeinflussen. Damit die physiologischen Funktionen des L-Plastin Signalweges in vivo getestet werden können und die Bedeutung von L-Plastin für verschiedene Erkrankungen in vivo analysiert werden kann, etablieren wir derzeit Mausmodelle, in denen die Expression von L-Plastin Zelltyp-spezifisch ausgeschaltet wird.